
|

|
Большая Советская Энциклопедия (цитаты)
|

|
|
|
|
Порфирины | Порфирины (далее П), широко распространенные в живой природе пигменты, в основе молекулы которых лежит порфин — структура из четырех колец пиррола
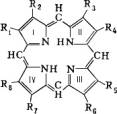
Порфирин:
R1=R2=R3=R4=R5=R6=R7=R8=
Протопорфирин:
R1=R3=R5=R8=3
R2=R4=—=2
R6=R7=24
Уропорфирин:
R1=R3=R5=R8=2
R2=R4= R6=R7=24
(см. формулу). Природные П различаются заместителями (R), среди которых наиболее распространены метильная (СН3), этильная (25), винильная (=) группы, остатки уксусной (2) и пропионовой (24) кислот. П обладают характерными спектрами поглощения и флуоресценции, которые служат для их идентификации. Наиболее биологически важны комплексы П с металлами и . Так, переносящие красные пигменты крови и мышц — гемоглобин и миоглобин содержат -порфириновый комплекс — гем. Аналогичные комплексы содержат цитохромы, играющие роль универсальных биохимических переносчиков электронов, а также ферменты каталаза и пероксидазы. Зеленые пигменты растений хлорофиллы — -комплексы П, витамин В12 (кобаламин) — Со-комплекс соединения, близкого к П Методом изотопных индикаторов показана общность путей биосинтеза П в клетках животных (гемоглобин) и растений ( началом которого служит конденсация глицина и янтарной кислоты (в форме сукцинилкофермента А) с образованием предшественника гема и — d-аминолевулиновой кислоты, а затем порфобилиногена и протопорфирина. П обнаружены также в выделениях животных — моче (уронорфирин), кале (копропорфирин), в скорлупе птичьих яиц, оперении птиц, раковинах моллюсков, а также в нефти, битумах и ископаемых органических остатках (часто в виде комплексов с и ). Абиогенное образование П связывают с путями эволюции. Нарушения обмена П (в т. ч. врожденные) приводят к заболеваниям человека — различным порфириям.
А. А. Красновский. |
Для поиска, наберите искомое слово (или его часть) в поле поиска
|
|
 |
 |
 |
|
|
|
Новости 22.06.2026 01:56:25
|
|
|
|
|
|
|
|
|
|
|


